
Ce texte a été publié à l'origine dans le n° 11 (1994) des Cahiers antispécistes. Il a été repris en 2018 dans l'ouvrage collectif La Révolution antispéciste.
Chapeau accompagnant l'article lors de sa parution dans les Cahiers antispécistes (1994):
Les espèces n’existent pas? Les antispécistes sont-ils devenus fous? Certes, depuis des années, les antiracistes, eux, affirment souvent que les races n’existent pas. On a bien des humains à peau claire ou foncée, à yeux bridés ou non, etc.; mais ceci n’en fait pas des races. Il s’agit simplement de traits individuels, souvent héréditaires et corrélés avec les populations mais ne permettant pas une telle subdivision de l’humanité.
La notion de race reste malgré cela bien vivace dans les esprits. Malgré les explications des antiracistes, basées sur des critères de scientificité, l’idée a du mal à prendre. L’existence des races paraît, aux racistes bien sûr mais pas seulement à eux, trop relever de l’expérience immédiate, d’une évidence naïve.
La notion d’espèce, au contraire, n’a pratiquement jamais été mise en doute par personne. Certes, les définitions successives qu’on en a donné au cours de l’histoire ont fait l’objet de controverses incessantes; mais chaque fois était en cause l’insuffisance d’une définition; jamais, du moins à ma connaissance, on n’a voulu jeter l’idée d’espèce elle-même aux orties. Jusqu’à très récemment à moi aussi m’aurait paru tout à fait farfelu de dire de l’espèce qu’elle est une notion aussi peu scientifique que celle de race. D’ailleurs, l’antispécisme n’a pas besoin de ça; qu’importent les catégories, scientifiques ou non, l’important c’est le fait qu’un être soit sensible ou non.
La même chose peut, cependant, se dire du racisme. Qu’importe que l’idée de race soit scientifique ou non; les catégories scientifiques ne sont pas en soi des catégories éthiques. Cependant, si les antiracistes ont cru bon de s’attaquer à la notion de race il devait bien y avoir une raison. L’idée en elle-même devait être porteuse d’un «quelque chose» de particulier. Or, historiquement et structurellement, les races correspondent à la tentative d’étendre plus avant les principes de la division linnéenne, laquelle, autrement, s’arrête à l’espèce. Il valait donc la peine d’aller voir de plus près si la classification linnéenne elle-même n’était pas porteuse d’un tel «quelque chose».
Les résultats de ma réflexion à ce sujet, que je vous livre ci-dessous, m’ont surpris moi-même. Je les crois pourtant bien fondés. Malgré les «évidences», naïves ou non.
62 min.
La science a besoin d'ordre: donc de classifications. C'est là la première justification qui peut venir à l'esprit, naïvement, quand on s'interroge sur la raison d'être de la notion d'espèce en biologie. Pour y voir clair, cette science pas plus qu'une autre ne peut se contenter d'envisager les objets qu'elle étudie – plantes ou animaux individuels – un par un; il lui faut pouvoir les insérer dans une structure et ainsi rapporter les faits particuliers à un ensemble plus général, et inversement pouvoir former des prédictions sur la base de la position d'un individu dans cet ensemble. Toutes les sciences font de même, dans leur domaine. Ainsi, par exemple, en géologie on connaît les roches métamorphiques, cristallines, sédimentaires et ainsi de suite. En astronomie, les étoiles sont classées selon leur type spectral, ou selon leur masse, ou selon leur appartenance à une séquence (naines, géantes, etc.). Cependant, on note deux traits très particuliers à la biologie sur ce sujet:
1. L'existence dans cette discipline d'un système de classification privilégié par rapport à tout autre, d'un caractère particulier (hiérarchique) et jouissant d'une réputation de «scientificité» spéciale;
2. Le fait qu'à la biologie en général et à cette classification tout particulièrement correspond un enjeu politique.
Détaillons:
1. Il existe en biologie une grille unique à laquelle est attribuée un statut privilégié de vérité, de scientificité. Il s'agit de la classification dite linnéenne1 ou, justement, «scientifique», qui découpe l'ensemble des individus vivants en catégories de base - ce sont aujourd'hui les espèces -, elles-mêmes regroupées en une hiérarchie de catégories supérieures emboîtées. Un individu fait partie d'une et d'une seule catégorie de base, et appartient aux autres, de niveau supérieur, seulement à travers la première. Je suis un être humain (mon espèce), et aussi un mammifère (ma classe), mais cette dernière catégorie n'est pas indépendante de la première, elle la contient entièrement. En somme, c'est en fonction de mon appartenance à une espèce donnée que je fais aussi partie d'autres catégories; je n'ai pas dans la hiérarchie linnéenne d'autres caractéristiques que celles qui découlent de mon espèce. Au contraire, par exemple, en astronomie, le fait qu'une étoile soit naine plutôt que géante ne dit pas si elle est de type spectral A plutôt que F – les quatre possibilités existent: naine A, naine F, géante A et géante F. La catégorie «étoile naine» traverse les frontières de la catégorie «type spectral A». Il n'existe aucune grille privilégiée de catégories de base ayant vocation à résumer la totalité de la position d'un individu. Il en est ainsi dans toutes les disciplines scientifiques ou autres; seule la biologie est dominée par une classification unique, totalitaire.
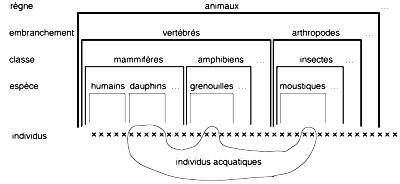
Bien sûr, dans chaque domaine particulier de la biologie, il y a aussi d'autres grilles de classification en usage. Par exemple, en écologie on distinguera animaux terrestres et animaux aquatiques; ces catégories traversent la frontière de celles de la classification linnéenne, y compris au niveau de l'espèce (on trouve à un moment donné des individus d'une même espèce de grenouilles sous forme de têtards, aquatiques, et d'autres sous forme adulte, terrestre; voir fig. 1). Mais ces autres grilles sont toujours perçues comme ad hoc, comme possédant un statut de vérité inférieur à la hiérarchie linnéenne, qui elle au contraire est scientifique, est la classification scientifique, la taxinomie2.
2. La classification biologique dite scientifique fonde à travers ses catégories de base une discrimination particulière, celle pratiquée par les humains à l'encontre des individus d'autres espèces. La biologie non seulement s'intéresse à la vie, mais aussi intéresse la vie, la nôtre; elle n'est pas neutre, encore moins que d'autres sciences. Au xixe siècle il semblait naturel de porter plus avant le système de classification linnéen, et de subdiviser les espèces à leur tour, en particulier l'espèce humaine. On distingua donc des races d'humains, et, fort commodément, on trouva que les frontières de ces nouvelles catégories de base coïncidaient avec celles de catégories politiques discriminatoires (celle des esclaves, par exemple). C'était alors la race, et non plus l'espèce, qui résumait tout ce que l'on pouvait dire d'un individu du point de vue de «la» taxinomie. À l'inverse, l'antiracisme actuel s'est pour une bonne part efforcé de dénoncer la notion de race, de la déclarer non scientifique. Pourtant, semblerait-il, toute catégorie est a priori scientifique, pour peu qu'elle repose sur des caractéristiques individuelles repérables; par exemple, la catégorie des individus à peau foncée («les Noirs») est parfaitement scientifique3, tout comme celle des droitiers. C'est donc non la scientificité en soi du fait de classer les individus humains, mais un quelque chose d'autre que possède la hiérarchie linnéenne, quelque chose auquel elle doit sa «scientificité» si particulière, qui était et qui reste un enjeu de débat politique.
D'ailleurs la notion de race, et généralement toute classification en dessous de l'espèce, ne peut s'intégrer de façon cohérente au schéma linnéen, qui exige que les individus transmettent à leurs descendants leur propre position taxinomique. La reproduction implique le plus souvent deux parents, et cette règle ne peut être satisfaite que s'ils occupent tous deux la même position. Or l'espèce, on le verra, est définie pour être la plus petite unité possible qui permette de conserver cette règle. Les sous-espèces, races et autres variétés n'arrivent jamais à accéder au prestige «scientifique» dont jouit la hiérarchie linnéenne4. On ressent cependant encore et toujours, malgré cette évidence, comme une envie chez de nombreux biologistes de prolonger la classification linnéenne en dessous de l'espèce, d'y intégrer toutes les variations génotypiques. La pierre de touche de la classification linnéenne, son atome, reste toutefois l'espèce. Il suffira pour l'instant de noter à nouveau que c'est aussi sur cet atome d'une classification non seulement réputée scientifique mais réputée la scientifique entre toutes que s'appuie la discrimination majeure opérée par les humains.
L'antiracisme a remis en question la scientificité de la notion de race humaine, insistant au contraire sur l'«unité» de notre espèce. Cette unité, cependant, est liée à des critères de toute évidence plus politiques ou éthiques que scientifiques. J'ai dit que cette remise en cause visait ainsi non la scientificité en soi mais un quelque chose de particulier attribué à la hiérarchie linnéenne, un quelque chose d'ordre en réalité idéologique. C'est qu'il y a effectivement – c'est le thème de cet article – un implicite dans la classification linnéenne, un implicite idéologique auquel elle doit son statut privilégié au sein des sciences biologiques.
Cet implicite, on n'a pas voulu le voir étendre à l'intérieur de l'espèce humaine parce qu'il en aurait menacé l'unité; au contraire, insiste-t-on, «tous les hommes5 naissent égaux». C'est-à-dire:
— égaux dans les faits; malgré leurs différences, il y aurait en chacun d'eux un quelque chose d'identique, une même essence; cette essence aurait un caractère factuel («tous les hommes naissent égaux» se présente non comme une prescription, mais comme un fait);
— égaux dans leurs droits, dans leur dignité etc., c'est-à-dire que de cette supposée identité factuelle naîtrait une prescription d'égalité de traitement, une égalité éthique. L'essence en question,«objet» métaphysique, aurait le pouvoir de déterminer une prescription.
Si le fait de subdiviser l'espèce humaine selon le système linnéen est capable d'en menacer l'unité d'essence et l'unité éthique, c'est bien que sous les catégories «purement scientifiques» de ce système, et particulièrement dans ses divisions atomiques, se cache une attribution d'essence. La classification linnéenne est perçue comme la classification parce qu'elle classe l'essentiel. C'est d'abord sous cet aspect que je vais l'examiner. On verra cependant ce faisant que si on lui ôte ce rôle de distributeur d'essences, elle n'est même plus une classification scientifique. Et son objet de base – l'espèce – non plus.
Ma critique s'articulera suivant les quatre sections ci-dessous:
I. La notion d'espèce ainsi que l'ensemble de la classification linnéenne et une bonne partie de l'appareil théorique correspondant ont été forgés avant la théorie de l'évolution, dans un cadre explicitement essentialiste.
II. La notion d'espèce, bien que privée officiellement de son fondement théorique essentialiste, est restée pratiquement identique dans sa substance depuis l'époque prédarwinienne. Néanmoins, sans le soutien de l'essence les espèces ne peuvent plus caractériser les individus, mais seulement leurs rapports. Le fait que la classification linnéenne reste vue comme la classification scientifique implique cependant que soit hypostasiée6 implicitement le concept d'espèce en une réalité caractérisant les individus, et correspond à la persistance sous-jacente de la vision essentialiste.
III. La classification linnéenne n'est pas une classification scientifique du tout, en ce sens qu'elle n'est pas théorique; son seul statut scientifique serait celui non de classification mais, sous sa forme «phylogénétique», d'hypothèse historique, c'est-à-dire portant sur l'évolution, testable (falsifiable), non conventionnelle. Il faudrait pour cela commencer à prendre la théorie de l'évolution au sérieux et en tirer les conséquences. La persistance de l'essentialisme implique la volonté au contraire de continuer à la considérer comme une classification, et comme la classification.
IV. La volonté de considérer une classification unique, hiérarchique, comme la classification scientifique est en soi totalitaire et tend de ce fait au racisme. Le racisme entre cependant en même temps en contradiction avec la logique de la classification linnéenne, qui veut que la position systématique se conserve d'une génération à l'autre, et qui ne peut donc accepter de classer différemment les individus quand ils sont interféconds. La systématique linnéenne se réfugie alors dans le spécisme, voyant tous les individus d'une même espèce comme essentiellement identiques. Mais le spécisme est à son tour en contradiction avec cette logique si on prend en compte l'évolution, ce que précisément la systématique rechigne à faire.
Au niveau des données historiques je m'inspirerai beaucoup du livre d'Ernst Mayr, Histoire de la biologie7. Mayr, biologiste et systématicien très reconnu et représentatif, se dit, comme tous les scientifiques actuels, opposé à la conception essentialiste de l'espèce. Nous verrons ce qu'il en est8.
I. Le cadre historique: l'essentialisme
Les bases de la théorie essentialiste de l'espèce
Si dans la notion moderne d'espèce l'idée d'essence est cachée – les essences n'ont plus leur place dans la science – cela n'a pas toujours été le cas. On n'a pas en fait à remonter bien loin pour trouver cette notion admise explicitement.
La vision du monde qui prévalait en Occident jusqu'à la fin du siècle dernier était créationniste. La terre avait été engendrée par un dieu environ quatre mille ans avant l'Ère chrétienne9, ensemble avec tous les êtres vivants «selon leur espèce10». Ces derniers, une fois créés, se reproduisaient, c'est-à-dire perpétuaient leur espèce malgré leur propre caractère éphémère.
L'espèce n'était pas indépendante des caractéristiques visibles des individus; c'était au contraire à travers elles qu'on l'appréhendait. Un cheval engendre un cheval, un chien un chien; et les chevaux, visiblement, diffèrent des chiens. Les espèces pouvaient ainsi se constater simplement, selon cette conception, comme regroupements d'individus semblables. Cependant cette similitude, qui se transmettait de génération en génération, était due à une chose impalpable et invisible, mais puissante, présente en chaque individu et capable de le déterminer tout entier, physiquement et moralement; une chose qui transcendait l'individu parce qu'il la partageait avec d'autres et parce qu'elle lui survivait; une chose immuable, créée à l'origine du monde par Dieu. Les individus, eux, étaient des expressions de cette chose, qui était leur essence. Ils n'étaient que les «représentants» de leur espèce (expression encore en usage aujourd'hui), de ce «moule intérieur» invariable que G.L. Buffon (1707-1788) rapportait au premier individu11 créé12:
Il y a dans la Nature un prototype général dans chaque espèce sur lequel chaque individu est modelé (...): le premier animal, le premier cheval par exemple, a été le modèle extérieur et le moule intérieur sur lequel tous les chevaux qui sont nés, tous ceux qui existent et tous ceux qui naîtront, ont été formés.
En chaque individu, il y avait donc ce moule intérieur, qui était soit strictement identique à celui d'un autre soit radicalement différent; les espèces étaient séparées par des «fossés infranchissables».
Le Plan de la Nature
Les espèces n'étaient cependant pas sans relations les unes avec les autres. On voyait bien, non seulement qu'il y avait des plantes et des animaux, mais aussi qu'un chat ressemble plus à un lion qu'à un poisson. Déjà Aristote avait distingué les animaux «avec sang» et «sans sang» – ces derniers correspondant grosso modo aux invertébrés actuels. La Nature, selon Aristote, ne faisant rien en vain, et ayant été, selon les chrétiens, créée par Dieu, il fallait découvrir le Plan de la Nature ou celui de Dieu au moment de la Création. Les espèces elles-mêmes devaient pouvoir être classées, en divisions et en subdivisions. Il ne s'agissait pas de déterminer un critère de classement adapté à une fin particulière pour ensuite départager les espèces en fonction, mais de chercher à redécouvrir un Plan préexistant, unique. Il y avait une et une seule systématique «vraie»; la recherche des critères était celle des critères «vrais». Les groupements retenus devaient être «vrais» – on dit aujourd'hui «naturels».
Ce point de vue, donc, est à l'origine de la systématique. Chaque division hiérarchique était fondée sur un critère précis; ainsi, la colonne vertébrale distingue les vertébrés des autres animaux; les mammifères sont les animaux à mamelles. Il fallait que le critère soit le vrai, soit celui du Plan de la Nature. Tous les critères ne pouvant pas être vrais en même temps, il fallait faire un choix, entre par exemple classer les chauve-souris, qui ont des ailes et des mamelles, soit avec les oiseaux, soit avec les mammifères. L'histoire de la systématique est celle des systèmes concurrents pour distinguer les bons critères. Chaque naturaliste avait le sien. Par exemple «Linné a donné à sa méthode de référence aux caractères floraux le nom (...) de 'système sexuel'. C'est qu'il accordait une importance primordiale à la reproduction. Elle indiquait, selon lui, les plans secrets du Créateur13.»
La théorie essentialiste face aux variations
Les individus n'étaient que des représentants imparfaits de leur essence puisqu'on voyait bien qu'il existait des variations au sein d'une même espèce (tous les chevaux ne sont pas identiques), alors que l'essence, elle, devait être immuable. «Tel père, tel fils»: c'est vrai seulement jusqu'à un certain point, malgré le fait que l'essence, responsable de la similitude des individus d'une espèce, se transmettait sans changements. Il en résultait une tension entre la théorie essentialiste et les faits observés qui peut se résumer ainsi:
— Les espèces sont un moule intérieur identique chez tous les individus qui le possèdent.
— Ce moule intérieur se traduit et se repère par l'aspect extérieur de l'individu.
— Cependant les individus d'une même espèce ne se ressemblent pas entièrement.
Le problème ne se posait pas qu'en théorie, mais aussi dans la pratique scientifique qui se développait. Comment savoir si deux individus ont ou non le même moule intérieur? On n'a directement accès qu'à leurs caractéristiques effectives, traduction imparfaite de leur essence; or le degré de ressemblance est difficile à mesurer objectivement, et par ailleurs les variations constatées, y compris au sein de la même espèce (entre père et fils par exemple), étaient parfois considérables. C'est pourquoi le botaniste anglais John Ray avait proposé en 1686 d'éliminer les caractères accidentels, c'est-à-dire non essentiels, pour déterminer les espèces14:
Après de longues recherches, il m'est apparu qu'il n'y avait pas de critère plus sûr pour déterminer les espèces que les traits distinctifs qui se perpétuent eux-mêmes par propagation à partir des graines. Ainsi, quelles que soient les variations se produisant chez les individus ou les espèces, si elles émanent des graines d'une seule et même plante, ce sont des variations accidentelles, et elles ne sont pas à considérer pour distinguer une espèce (...).
Les caractères essentiels sont tous ceux qui se transmettent. Interprété littéralement, ce critère fait de tout trait héréditaire un caractère spécifique – dans le contexte de nos connaissances actuelles qui nous apprennent que chaque individu est le plus souvent génétiquement unique, il y aurait alors presque autant d'espèces que d'individus. On y reviendra.
Buffon ci-dessus à propos du premier individu de chaque espèce parle de prototype, c'est-à-dire de premier type. Puisqu'il s'agissait dans la théorie en question d'un individu réel, il devait contenir toutes les caractéristiques d'un organisme entier. Le type avait un certain nombre de pattes, une certaine couleur, et ainsi de suite. Un individu peut s'écarter de son type, en cela il sera atypique. Si une mouche a cinq pattes, alors que normalement elle en a six – si c'est une mouche estropiée, par exemple, c'est-à-dire ayant subi un accident – ce caractère sera vu comme accidentel, comme un écart au type. Il s'agit là de la théorie typologique de l'espèce, c'est-à-dire de la variante de la théorie essentialiste dans laquelle l'essence contient de quoi déterminer l'individu entier. On devait pouvoir trouver dans la nature des individus presque parfaits, particulièrement fidèles à leur moule intérieur; quand un naturaliste en prenait un, il le prélevait et le conservait (mort), le décrivait par le menu et le baptisait type de l'espèce, en souvenir du type primordial, cet ancêtre originel à qui il ressemblait si bien. Il n'y a qu'un seul mètre étalon dans le monde, mais on trouve répertoriés dans les instituts scientifiques actuels quelques millions de «types», plantes séchées et animaux formolisés (lesquels ne demandaient pas tant d'honneur) constituant encore aujourd'hui, pour chaque espèce, la référence officielle.
Des choix douloureux pour l'essentialisme
La conception typologique de l'espèce s'appliquait cependant mal dans certains cas. Chez les chats, par exemple, il n'y a pas de couleur particulière nettement plus fréquente qu'une autre. Qu'un chat fût noir, blanc ou tigré, il paraissait arbitraire d'y voir un écart au type. Selon la conception typologique, pourtant, il devait bien y avoir eu un premier chat, donc un type, avec une couleur bien précise. Il y avait là une difficulté pour la conception typologique de l'espèce, mais non pour la conception essentialiste en général. Il suffisait de considérer que l'essence ne contient pas de quoi déterminer un individu entier; que certains caractères sont libres, ou du moins ne sont pas entièrement spécifiés par l'essence, laquelle contiendra par exemple dans le cas du chat seulement une gamme de couleurs possibles (noir, blanc, tigré, mais pas vert). Dans la pratique systématique d'aujourd'hui on s'en sort souvent en conservant comme type de référence non pas un individu mais plusieurs, censés couvrir la gamme des possibilités laissées indéterminées par l'espèce.
Les variations restaient cependant problématiques y compris pour l'essentialisme lui-même quand elles étaient héréditaires. J'ai noté ce problème à propos de la caractérisation que donnait Ray des variations accidentelles. Si la couleur du chat est accidentelle, c'est-à-dire n'est pas déterminée par son essence, comment rendre compte du fait d'observation selon lequel elle est tout de même héréditaire? On connaissait bien les races de chevaux, de chiens, et ainsi de suite. Puisque l'idée d'essence servait à expliquer l'hérédité – le fait qu'un cheval engendre un cheval, etc. – il y avait comme un vide explicatif si elle ne déterminait pas tous les caractères héréditaires – si elle n'expliquait pas qu'un cheval blanc engendre un cheval blanc. Si au contraire l'essence était porteuse de tous les caractères héréditaires, il fallait élever au rang d'espèce toute race rencontrée. Et de fait, certains auteurs du xixe siècle faisaient de chaque variant trouvé une espèce nouvelle15. Le même schéma a conduit à une floraison de races humaines – noire et blanche bien sûr, mais aussi nordique, latine, etc. – avec chaque fois une essence différente. On se heurtait cependant à nouveau à deux problèmes, qui menaçaient de détruire l'édifice entier. D'abord, ces nouvelles espèces étaient interfécondes: les chevaux arabes se reproduisent très bien avec les chevaux européens. Quid de l'essence de la progéniture? On la baptisait «sang-mêlé», c'est-à-dire qu'on supputait la cohabitation en un seul être de deux essences. Il s'agit là d'une opération métaphysique délicate et une bonne part des efforts du racisme visaient à suppléer aux insuffisances des lois de la nature par des lois humaines interdisant l'apparition de tels «monstres» (cf. la volonté nazie de pureté raciale). Ensuite, en raison aussi de cette interfécondité, les frontières effectivement constatées dans la nature manquaient souvent complètement de netteté, et les catégories repérées n'étaient souvent pas univoques, c'est-à-dire que selon une certaine classification, a pouvait se retrouver classé avec b, et selon une autre, avec c. Il fallait subdiviser toujours plus, mais alors tout finissait par se mêler et le concept d'essence immuable et héréditaire perdait toute vraisemblance.
Pour sauvegarder l'édifice essentialiste, et malgré les envies de subdivision à l'infini toujours présentes en raison de la logique de l'équation essence = hérédité, on était bien obligé de s'en tenir à la caractérisation des espèces énoncée en 1749 par Buffon16:
On doit regarder comme la même espèce celle qui, au moyen de la copulation, se perpétue et conserve la similitude de cette espèce, et comme des espèces différentes celles qui, par les mêmes moyens, ne peuvent rien produire ensemble.
C'est-à-dire que les races ne sont pas des espèces; l'espèce inclut uniformément tous les individus qui peuvent se reproduire ensemble. Certes, dans ce cas, l'essence ne spécifie pas un ensemble complet de caractères, ni même l'ensemble des caractères héritables; elle n'expliquait plus l'hérédité, du moins pas dans tous les cas, mais on pouvait malgré tout continuer à croire en son existence; et de fait c'est ce qu'on a fait, jusqu'au début de ce siècle. L'édifice essentialiste restait entier, même s'il lui manquait une bonne partie de ses fondements. Au xxe siècle, on a trouvé pour l'hérédité des explications indépendantes de l'essence et plus vraisemblables (le matériel génétique); l'idée d'essence, privée du coup de ses dernières raisons d'être explicatives, contredite par la théorie de l'évolution, aurait dû disparaître. Aurait dû: en réalité, elle a continué à subsister, non plus ouvertement, mais, comme je l'expliquerai plus loin, de façon implicite dissimulée dans la structure de la pyramide linnéenne.
La théorie essentialiste jouissait au moins d'une cohérence interne. Elle permettait de fonder une classification authentique en tant que telle. Les essences étaient supposées réelles; l'essence «chat» existait – identifiée ou non au premier chat – et existerait encore conceptuellement même s'il n'y avait plus aucun chat au monde. La théorie, c'était l'existence des essences; la classification consistait à situer les individus par rapport à ces catégories données a priori. Si un individu avait l'essence «chat», on le classait dans la catégorie chat. Il s'agissait cependant d'une classification d'un genre particulier, en ce que la catégorie des objets étudiés était justement leur essence, et donc épuisait la totalité de ce qu'on pouvait en dire – hormis ce qui relevait de l'accidentel, c'est-à-dire possédait un statut de réalité moindre. Ainsi la classification linnéenne, fondée sur les essences, était-elle non seulement une classification scientifique, mais la classification scientifique.
La pierre de touche du repérage des espèces dans la caractérisation par John Ray citée plus haut, c'est la reproduction. Georges Cuvier (1769-1832) insistait encore là-dessus17:
Nous estimons qu'une espèce est représentée par la descendance totale du premier couple crée par Dieu (...). Quel moyen avons-nous de nos jours pour redécouvrir la trace de cette généalogie? Ce n'est assurément pas dans la ressemblance structurale. Il ne reste en réalité que la reproduction et je soutiens que c'est le seul caractère certain et infaillible qui permette la reconnaissance des espèces.
La reproduction, caractère relationnel (relation entre deux individus), n'intervenait cependant pas en tant que critère de définition; il s'agissait, comme le dit ici Cuvier, d'un moyen de découvrir, de reconnaître les espèces, lesquelles préexistaient à l'investigation scientifique. L'espèce elle-même, bien que repérée au moyen de la reproduction, restait bel et bien une caractéristique individuelle, une caractéristique que chaque individu possédait indépendamment des autres.
Je me suis étendu longuement sur le concept essentialiste d'espèce, alors qu'on pourrait croire qu'aujourd'hui, la théorie de la création divine ayant enfin cédé la place à la celle de l'évolution, son seul intérêt resterait historique. On pourrait s'attendre en particulier à ce que la notion d'espèce ait radicalement changé depuis la publication par Darwin en 1858 de L'Origine des espèces; voire, puisque l'idée d'espèce était entièrement dépendante de celle d'essence, on pourrait s'attendre à ce que la seconde ait entraîné la première dans sa chute, que les espèces aient disparu de la biologie ou tout du moins aient été remplacées par un autre concept dépourvu de la prééminence qui revient à une notion censée traduire directement une entité transcendante. On verra qu'il n'en est rien. L'espèce était et demeure la pierre de touche de la systématique. Son critère de reconnaissance est encore la reproduction. La volonté de découper les espèces à l'infini et de baptiser autant de sous-espèces que possible est toujours bien vivante. Toute nouvelle espèce découverte doit être accompagnée de son type et recevra en retour son nom de baptême, un beau binôme linnéen. Et l'existence des variations, matière première du darwinisme, reste le cauchemar des systématiciens.
La réalité, on va le voir, est que la notion d'espèce actuellement admise par la grande majorité des scientifiques, appelée «concept biologique de l'espèce», a été forgée sous la simple pression des faits, indépendamment de Darwin et de toute remise en cause de l'essentialisme. On peut dire qu'elle a été modifiée le moins possible dans sa définition par rapport aux conceptions antérieures et qu'elle est restée parfaitement compatible avec l'essentialisme; voire, elle ne peut se comprendre comme théorie de classification que dans un cadre essentialiste, lequel n'est rejeté qu'en apparence.
Au niveau des groupes taxinomiques supérieurs à l'espèce, il y a eu là aussi bien peu de changements. Bien qu'aujourd'hui on ne parle plus ouvertement de Plan de la Nature, les groupes reconnus dans la systématique, y compris les nouveaux, sont encore très généralement nommés en fonction d'un unique critère «vrai18». On a bien été obligé cependant de reconnaître que souvent un caractère même vrai pouvait faire défaut; par exemple, si en botanique la présence de chlorophylle définit le grand groupe des chlorophylliens, il existe des plantes dont les ancêtres ont perdu la fonction chlorophyllienne; elles restent classées parmi les chlorophylliens, elles sont chlorophylliennes sans l'être. On qualifie la perte de chlorophylle de secondaire; la plante reste chlorophyllienne dans l'âme.
Devant ce genre de difficultés, qui se posent à un niveau pratique aigu dans de nombreux groupes au sein desquels les similitudes s'entrecroisent et qu'on ne sait vraiment pas comment classer, la systématique d'aujourd'hui se tourne vers la méthode dite «phénétique», basée sur le traitement, par ordinateur, des ensembles de similitudes et de différences dans le but de dégager, encore et toujours, la classification vraie. Les dichotomies ne se basent plus sur un unique caractère «vrai»; ce qui le remplace, c'est l' «objectivité» des chiffres (dopée par la présence de l'ordinateur), le but étant resté le même. Mais quel sens cela peut-il avoir de chercher la classification vraie s'il n'y a pas un Plan de la Nature? On pourrait répondre que l'existence des groupes taxinomiques est aujourd'hui mise en relation avec l'histoire évolutive et que la recherche taxinomique vise à la redécouvrir. C'est là – j'en reparlerai – le point de vue phylogénétique19, que justement les systématiciens phénéticiens récusent explicitement. Ils veulent leur méthode «objective», indépendante de toute hypothèse (terme auquel ils donnent une connotation péjorative) sur la phylogenèse des espèces à classer. La volonté de dégager une classification unique n'a plus alors d'explication, à moins qu'on ne suppose que l'idée de Plan de la Nature est encore déterminante dans les structures mêmes de leur pensée.
La systématique est restée la systématique, et l'est restée sous sa forme linnéenne. Les débats à son propos sont toujours aussi vifs. On se torture encore la cervelle pour décider si oui ou non une unité taxinomique est «naturelle». Un taxinomiste du début du siècle dernier serait peu dépaysé s'il renaissait aujourd'hui dans un congrès de systématique; pour comprendre le fond des débats de la taxinomie moderne, il vaut bien mieux croire en la Création divine qu'en l'évolution darwinienne.
II. Le concept actuel d'espèce
Le concept moderne, dit «biologique», de l'espèce est, nous dit Ernst Mayr, en rupture totale avec le passé20:
(a) Le vieux concept d'espèce, basé sur l'idée métaphysique de l'essence, est si différent du concept biologique d'une population reproductivement isolée, que le passage graduel de l'un à l'autre fut impossible.
Mais pourtant: on trouve dès le paragraphe suivant21:
(b) Pour un biologiste moderne, il n'y a vraiment pas loin de la définition de Ray, exprimant un essentialisme modéré – «une espèce est l'ensemble de tous les variants qui sont potentiellement les rejetons des mêmes parents» – à la définition de l'espèce basée sur le concept des seules communautés reproductives. Encore plus proche était la définition de Buffon: «Une espèce est une succession constante d'individus similaires qui peuvent se reproduire ensemble», et dont les hybrides sont stériles.
Ray, pas plus que Buffon, n'était comme dit ici Mayr un essentialiste «modéré» – il était, comme tous ceux de son époque, un essentialiste tout court. Il n'existait pas alors d'autre conception de l'espèce que celle basée sur l'essence. La caractérisation de l'espèce par Ray que cite Mayr en (b) est parfaitement compatible avec l'essentialisme, tout comme celle de Buffon. Mayr parle ici de définitions; on a vu cependant, dans la citation que je donne plus haut de Buffon, que pour celui-ci l'espèce était déjà définie, comme essence («moule intérieur») et n'avait donc pas besoin d'autre définition; la phrase de lui que cite Mayr en (b) est donc non une définition mais seulement une technique de reconnaissance. Il est donc normal qu'on n'y parle pas explicitement d'essences. Il ressort du passage (b) de Mayr que le concept moderne d'espèce est presque identique en substance à la conception essentialiste; mais le passage (a) du même auteur nous dit exactement le contraire. Qu'en penser?
Mayr parle ci-dessus de l'espèce moderne comme «concept biologique d'une population reproductivement isolée», et plus loin comme défini sur la base du «concept des seules communautés reproductives». Nous allons voir comment, à partir de l'énoncé des critères déjà utilisées par les essentialistes pour reconnaître les espèces, on aboutit, sans faire aucunement intervenir l'évolution, à l'énoncé utilisé aujourd'hui pour caractériser cette notion.
Partons de l'énoncé de Buffon déjà reproduit ci-dessus:
On doit regarder comme la même espèce celle qui, au moyen de la copulation, se perpétue et conserve la similitude de cette espèce, et comme des espèces différentes celles qui, par les mêmes moyens, ne peuvent rien produire ensemble.
La précision «et conserve la similitude de cette espèce» n'a pas en fait un rôle de critère, puisqu'elle était supposée toujours vérifiée; elle n'est là que pour préciser la pensée, et l'auteur l'a omise dans la réciproque. On peut donc paraphraser cet énoncé ainsi:
Deux espèces sont en fait une seule si et seulement si elles sont interfécondes22.
L'énoncé semble pécher par circularité, puisqu'il faut partir de deux espèces pour en trouver une. On peut néanmoins prendre comme unité de base des groupes d'individus proches parents, comme étant par principe de la même espèce (on ne peut partir simplement des individus, puisque deux mâles, par exemple, ne sont pas interféconds); on obtient alors:
(i) Un individu a est de la même espèce qu'un autre b s'il en est un proche parent ou si a, ou un proche parent de a, est susceptible de se reproduire avec b, ou un proche parent de b23.
Pour Buffon, nous l'avons vu, ce genre d'énoncé n'avait pas un statut de définition, mais de méthode de repérage d'entités – les espèces – préexistantes. Je vais néanmoins montrer comment on peut vouloir faire de cet énoncé une définition, et qu'on arrive ainsi à un concept d'espèce identique au concept moderne, sans nullement faire référence à la théorie de l'évolution.
En tant que définition, l'énoncé (i) ne définit pas d'emblée l'espèce d'un individu, mais la relation binaire est-de-la-même-espèce-que, à lire comme un seul mot, un mot nouveau puisqu'on le définit; comme si on disait, par exemple «a droze b si et seulement si...». Cependant le but n'est pas de prendre la relation comme un tout indécomposable; c'est bien aux espèces qu'on s'intéresse. Comment le fait de définir est-de-la-même-espèce-que pourrait-il définir le mot «espèce»?
Une méthode existe pour extraire un concept de catégorie d'une telle expression. Elle se base sur les propriétés des relations mathématiques dites d'équivalence, c'est-à-dire possédant les propriétés de réflexivité, de symétrie et de transitivité. Ainsi une relation – nommons-la justement «droze» – est d'équivalence si:
— elle est réflexive: pour tout individu a, on a a droze a;
— elle est symétrique: chaque fois que a droze b, on b aussi b droze a;
— elle est transitive: quand a droze b et b droze c, alors a droze c.
Ces trois propriétés doivent être satisfaites si on veut pouvoir remplacer le mot «droze» par l'expression «est de la même espèce que», lue en la décomposant mot par mot. Réciproquement, on montre que quand une relation est d'équivalence, elle permet de découper l'ensemble des individus en classes d'équivalence et devient alors synonyme de «est de la même classe d'équivalence que». Définir une relation permet ainsi, quand elle est d'équivalence, de définir du même coup des classes d'équivalence, et de remplacer la relation entre deux individus par une relation entre un individu et sa classe. Il suffit alors en quelque sorte d'hypostasier la classe, de l'imaginer comme une réalité indépendante des éléments dont elle est composée, pour croire être arrivé à une classification.
La relation est-de-la-même-espèce-que est-elle donc d'équivalence? Telle que définie par l'énoncé (i), elle est réflexive (un individu étant toujours prochement apparenté à lui-même), et symétrique (par exemple «x se reproduit avec y» est la même chose que «y se reproduit avec x»). Par contre la transitivité pose problème. Rien ne permet a priori de dire que si a (ou un de ses proches parents) peut se reproduire avec b (ou un de ses proches parents), et qu'il en est de même pour b et c, alors il en est de même aussi pour a et c. Et de fait des contre-exemples existent; on les appelle parfois «espèces en anneau brisé». Voici le cas des goélands bruns et des goélands argentés tel que présenté par Richard Dawkins24:
En Grande Bretagne il s'agit de deux espèces nettement distinctes, tout à fait différentes en couleur. N'importe qui peut les distinguer. Mais si on suit la population de goélands argentés vers l'ouest pour contourner le Pôle nord, en traversant d'abord l'Amérique du Nord, puis l'Alaska et la Sibérie pour revenir finalement en Europe, on remarquera un fait curieux. Progressivement, les «goélands argentés» ressemblent de moins en moins à des goélands argentés et de plus en plus à des goélands bruns, et il s'avère en fin de compte que nos goélands bruns européens forment en fait l'autre extrémité d'un anneau qui débute avec des goélands argentés. À chaque point de l'anneau, les oiseaux ressemblent suffisamment à leurs voisins pour pouvoir se reproduire avec eux; ceci, du moins, partout ailleurs qu'aux deux bouts de l'anneau, en Europe. En ce point, les goélands argentés ne se reproduisent jamais avec les goélands bruns, bien qu'ils leur soient liés par une série continue de collègues interféconds tout autour du monde.
Est-ce qu'un goéland argenté est-de-la-même-espèce-que un goéland brun? Certainement non, si on se réfère à l'énoncé (i); cependant il est-de-la-même-espèce-que un goéland américain, qui lui-même est-de-la-même-espèce-que un goéland alaskien, et ainsi de suite jusqu'au goéland brun. Parce que la relation n'est pas transitive, elle ne peut servir à définir une classe d'équivalence, qui serait l'espèce; on ne peut la décomposer comme voulant dire «est de la même espèce que», sans les tirets. Si on le faisait, par comparaison directe le goéland argenté et le goéland brun seraient d'espèces différentes, et par comparaison indirecte de même espèce; d'où contradiction.
La solution semble ne pouvoir être que de considérer malgré tout ces deux types de goélands comme une seule espèce, ensemble avec tous les types intermédiaires. Il faut pour cela modifier la définition (i), en pratiquant ce qu'on appelle sa fermeture transitive:
(ii) Un individu est-de-la-même-espèce-que un autre si la relation définie en (i) se vérifie directement, ou si elle se vérifie indirectement, à travers une chaîne d'intermédiaires.
Cela revient en substance au même que de dire:
(iii) Un individu est-de-la-même-espèce-que un autre s'il est susceptible d'avoir dans l'avenir un descendant commun avec lui.
En effet, si le goéland argenté ne s'accouple pas avec le goéland brun, il est cependant susceptible de le faire avec le goéland alaskien; ses descendants successifs peuvent de proche en proche faire le tour du pôle et il pourra se retrouver avec un descendant en commun avec un goéland brun.
Il s'agit alors bien d'une relation d'équivalence, c'est-à-dire réflexive, symétrique et transitive, et on peut, comme je l'ai dit, passer de la définition de la relation à celle des classes d'équivalence elles-mêmes, c'est-à-dire extraire de la relation est-de-la-même-espèce-que la notion d'espèce. L'espèce d'un individu, ce sera l'ensemble des individus avec lesquels cette relation est vérifiée, c'est-à-dire avec lesquels il est susceptible d'avoir un descendant commun. L'espèce d'un goéland brun sera l'ensemble des goélands de l'anneau, sa «communauté reproductive», «reproductivement isolée» de l'extérieur et sans frontières reproductives internes. On retrouve ainsi la notion moderne d'espèce telle que caractérisée ci-dessus par Mayr.
Comme je l'ai annoncé, pour aboutir à la définition moderne des espèces en partant du critère de reconnaissance énoncé par Buffon, on n'a nullement eu besoin de faire intervenir l'évolution. Je n'ai pas non plus fait intervenir dans cette transformation la notion d'essence, mais rien n'y est contradictoire avec celle-ci. Buffon pouvait aboutir, voire devait aboutir s'il prenait en compte l'existence d'espèces en anneau brisé, à la même notion sans aucunement renier l'essentialisme; la seule différence étant qu'il s'agirait pour lui d'une caractérisation et non d'une définition. La fermeture transitive introduit des cas dans lesquels deux individus de même essence ne se reproduisent pas entre eux; mais il n'y a là aucune difficulté théorique supplémentaire, des raisons «accidentelles» pouvant par exemple empêcher deux individus de s'accoupler25. Buffon aurait reconnu les mêmes espèces.
Il est étrange que dans deux cadres théoriques si différents, l'un essentialiste et l'autre officiellement évolutionniste, on reconnaisse les mêmes catégories. Nous allons voir cependant que la définition moderne de l'espèce est implicitement essentialiste, parce que, justement, elle se veut la définition de quelque chose de l'individu. Il ne suffit pas en effet, pour obtenir une classification scientifique, de pouvoir attribuer à chaque objet une catégorie; il faut aussi que cette attribution puisse se faire de façon non arbitraire, selon un critère relatif à l'individu seul. Or l'espèce définie comme classe d'équivalence selon la relation (iii) n'est pas un objet théorique; c'est un objet du monde réel, qui intègre l'ensemble des individus qui y vivent. Qu'un nouveau chat naisse, et l'ensemble des individus avec lesquels mon chat Ek pourrait se reproduire change; son espèce change. L'espèce d'Ek ne dépend pas que des caractéristiques d'Ek; elle dépend de circonstances qui n'ont rien à voir avec Ek elle-même. Pire, le fait que a soit de la même espèce que b ne dépend pas seulement de a et de b; l'introduction de la fermeture transitive a transformé une relation qui était binaire au départ en une affaire à trois ou plus. C'est pourtant la volonté de voir en la classification linnéenne une classification scientifique, la classification scientifique, qui implique malgré cela de considérer qu'Ek est un chat, c'est-à-dire d'hypostasier la notion d'espèce pour en faire la catégorie scientifique de l'individu. L'idée existe donc implicitement que les espèces sont autre chose qu'un ensemble d'individus; c'est-à-dire qu'elles existent à un niveau théorique. Comme nulle part pourtant dans la théorie on ne trouve de tels objets26, c'est bien que sans le dire on fait référence encore à la vieille idée d'essence.
Le seul élément de la théorie essentialiste qu'on ne retrouve pas dans le concept actuel d'espèce, c'est le type, qui n'en était pas un élément obligatoire. C'était seulement si l'essence contenait un ensemble complet de caractères qu'il s'identifiait à un type. À partir d'une classe d'équivalence définie par la relation (iii) on ne retrouve pas un type; les types sont malgré cela encore omniprésent dans la systématique contemporaine. Ils le sont dans le langage courant aussi, comme en témoigne l'idée que par exemple un chat puisse ne pas être un «vrai» chat («Mon chat ne chasse jamais, ce n'est pas un vrai chat.»).
Le fait que dans l'esprit des biologistes actuels le «concept biologique d'espèce» ne correspond pas à une vraie définition a pour conséquence qu'ils ne cessent pas d'en changer. Mayr, par exemple, proposait en 1942 celle-ci: «Les espèces sont des groupes de populations dont les membres peuvent se croiser entre eux, réellement ou potentiellement, et qui sont reproductivement isolés d'autres groupes27.» Plus loin28, il envisage la question de la reproduction asexuée29. L'application de sa définition de 1942 conduirait, dit-il, au fait «absurde de considérer [chaque individu] comme une espèce séparée». Absurde pourquoi? La définition existe, elle s'applique; son résultat ne peut être absurde que si on le veut conforme à idée d'espèce préexistante, indépendante de la définition exhibée. L'idée d'avoir une espèce par individu est effectivement absurde dans le cadre de la pensée essentialiste. Pour s'en sortir, pour faire mieux coller la «définition» qu'il donne à ce quelque chose qu'il ne nomme pas, il y ajoute la notion de niche écologique. Ainsi peut-elle «mieux rendre compte de la plupart des situations30». Chez les bactéries se pose le problème inverse: leur sexualité transgressive ferait réunir dans une même espèce des organismes trop différents. On pourrait s'attendre à ce que Mayr en conclue simplement que le concept d'espèce s'avère inutile chez les bactéries. On peut très bien les étudier sans cela. Mais non: le concept d'espèce est un must, c'est la vérité d'un organisme. Il faut donc continuer les recherches afin de trouver, pour elles aussi, le «concept d'espèce qui [leur] convient le mieux31.»
Les biologistes ont décidément du mal à se défaire de l'essentialisme – comme l'ensemble de la population d'ailleurs, mais c'est là un autre problème. Mayr lui-même nous cite cet extrait de L'Origine des espèces: «Lorsque les vues exposées dans cet ouvrage (...) seront admises par tout le monde (...), les systématiciens ne seront plus sans cesse hantés par le doute obscur de savoir si telle ou telle forme est, dans son essence, une espèce. Cela, j'en suis sûr, et j'en parle par expérience, ne sera pas un mince soulagement32.» Il faut croire que ce jour n'est pas encore venu.
III. Le statut scientifique de la classification linnéenne
Une classification consiste à choisir des critères et à classer les objets selon la manière dont individuellement ils y répondent. Au niveau du choix des critères à utiliser on tient compte évidemment de l'ensemble de la population à étudier; si tous les humains mourraient avant quarante ans, on ne prendrait pas la peine de distinguer la catégorie «humains de moins de quarante ans». Mais les critères une fois choisis s'appliquent à chaque individu indépendamment de la population; le fait que j'aie ou non moins de quarante ans s'applique à moi, et resterait tout aussi vrai ou faux si j'étais le seul humain sur terre.
On a vu qu'il en va tout autrement de la classification linnéenne. L'espèce n'est définie que par rapport à la population. Il en est de même aux niveaux plus élevés, pour le genre, la famille, etc.; cela est particulièrement apparent dans la méthode dite «phénétique», basée mécaniquement sur la «distance» entre les caractères des espèces différentes pour dire si oui ou non on doit les ranger dans un même genre, et ainsi de suite jusqu'au sommet de la hiérarchie. Puisqu'à chaque étape on classe les objets (individus, espèces, genres, etc.) non en fonction de leurs caractéristiques mais de leurs relations, on peut dire que ce n'est pas en fait une classification du tout; à moins de considérer que, comme je l'ai dit, il s'agit en fait non de classer mais de redécouvrir par cette méthode le «Plan caché du Créateur».
À la méthode phénétique s'oppose chez les systématiciens celle dite phylogénétique, appelée aussi cladisme en 1950 par son inventeur, W. Hennig. Cette fois il s'agit de baser la systématique entièrement sur l'évolution, et plus du tout sur les caractères effectifs des individus. La hiérarchie linnéenne est alors vue comme un simple reflet de la généalogie. Il ne s'agit cependant en fait plus d'une classification du tout: il s'agit d'un historique. L'ensemble des mammifères par exemple est dans cette hiérarchie un groupe correct (monophylétique) si, et seulement si, il regroupe tous les descendants du plus proche ancêtre commun33. À la place de la hiérarchie linnéenne on a alors un ensemble d'hypothèses scientifiques portant sur un objet bien précis, l'histoire évolutive, et qui sont en principe testables, falsifiables. On ne peut décider cette hiérarchie par convention, on ne peut que poser des hypothèses et chercher à les tester. En particulier, dans les cas où on ne dispose pas pas des données paléontologiques nécessaires, on ne peut établir la «classification». Ceci est une situation courante dans tout domaine scientifique, et il n'y a pas lieu de s'en émouvoir; pourtant, pour la plupart des biologistes, elle est intolérable. Il leur faut leur classification, il leur faut la classification systématique. C'est sans doute cette pression qui a conduit de nombreux «continuateurs» du cladisme à abandonner ce qui représentait le fondement même de leur théorie, à savoir le fait qu'elle devait traduire l'évolution, et à revenir à une méthode qui, à l'instar de la méthode phénétique, cherche à dégager le «Plan de la Nature» à partir de la comparaison des caractères. La prise au sérieux de l'évolution se montrait une fois de plus incompatible avec les fondements de la systématique linnéenne.

Un des reproches majeurs faits à la méthode phylogénétique était de regrouper ce qu'on ne voulait pas mettre ensemble. Par exemple la généalogie des reptiles et des oiseaux se présente comme dans la figure 2. On pense en effet que les oiseaux descendent d'un reptile particulier B qui a aussi donné naissance aux crocodiles. Le groupe contenant les oiseaux, les crocodiles et les autres reptiles a pour plus proche ancêtre commun A. Il contient aussi tous les descendants de A. Il s'agit donc d'un groupe monophylétique. Mais si le groupe des reptiles seuls – crocodiles et autres – a lui aussi le même plus proche ancêtre commun A, il lui manque pour être monophylétique les oiseaux, qui eux aussi descendent de A. La systématique traditionnelle, pourtant, établit le groupe des reptiles sur la base de leur similitude; les oiseaux, à l'inverse, ont beaucoup évolué depuis leur divergence d'avec B. On répugne à les classer ensemble avec les reptiles, cela ne semble pas correspondre au «Plan de la Nature». On veut pouvoir dire que les crocodiles sont des reptiles, qu'ils ont un quelque chose d'ontologique en commun avec les autres reptiles et que les oiseaux n'ont pas. On veut une hiérarchie linnéenne qui reflète cette vérité. On reproche donc à la méthode phylogénétique de ne pas intégrer le degré de divergence des groupes. Si pourtant on accepte cette méthode pour ce qu'elle est, on n'a pas à lui demander autre chose que de refléter l'histoire évolutive. Elle ne nie pas le fait que les oiseaux aient pu beaucoup changer depuis leur divergence d'avec les crocodiles; ce n'est tout simplement pas son sujet.
Pour permettre à la hiérarchie linnéenne de continuer à refléter la vérité des êtres, sans pour autant faire totalement l'impasse sur l'évolution, la solution adoptée aujourd'hui par la plus grande partie de la communauté scientifique s'appelle «méthode évolutionniste». Il s'agit en fait de sauvegarder l'esprit de la classification linnéenne en y intégrant l'évolution comme s'il s'agissait d'un caractère de plus, dont il faudrait tenir compte aussi. Mayr, qui la défend, accepte pour elle l'appellation d'«éclectique» parce qu'elle fait feu de tout bois. Elle ne possède aucun statut théorique clair; cette absence traduit tout simplement la présence cachée d'un objet théorique sous-jacent, le fameux «Plan de la Nature», pour la recherche duquel tout indice est bon.
La systématique linnéenne a décidément du mal à digérer Darwin. Et de fait: le créationnisme moderne, qui sévit encore puissamment dans certains pays, a beau jeu de profiter de ces contradictions. Il lui suffit d'interpréter la systématique linnéenne à la lettre. Chaque individu a son espèce; il est un humain, ou il est un singe. L'espèce se transmet par la reproduction; un singe ne peut donc engendrer un humain. La réponse typique du biologiste d'aujourd'hui sera de dire qu'il ne faut justement pas prendre la systématique à la lettre. Mais alors comment faut-il la prendre? Qu'est-ce alors exactement que l'espèce?
IV. Systématique linnéenne, racisme et spécisme
La volonté de considérer une classification unique, hiérarchique, comme la classification scientifique est en soi totalitaire. Prise à la lettre, elle signifie que du point de vue scientifique, donc du point de vue de la réalité vraie, nous sommes une seule chose, notre espèce. La catégorie de base où je suis rangé résume la totalité de ce que l'on peut, scientifiquement, dire de moi. Si on connaît mon espèce on connaît aussi mon genre, ma famille, mon ordre, ma classe et mon embranchement.
Du fait de ce que les humains sont pourtant bien divers, et que souvent leurs différences, loin de sembler accidentelles, se transmettent héréditairement, la classification linnéenne a du mal à s'arrêter à l'espèce. Si elle doit résumer tout ce qui est essentiel, et que les humains sont différents, il lui faut subdiviser leur espèce. Il lui faut distinguer les races, attribuer à chacune son essence. Un Noir sera un humain, et ensuite un mammifère, mais il sera d'abord un Noir. Tout comme cette logique n'admet aucun groupe contenant seulement certains chats et certains chiens – un tel groupe «n'existe pas», n'est pas une «catégorie scientifique», un «groupe naturel» – elle ne peut admettre d'attribuer un statut de pleine réalité à un groupe contenant seulement certains Noirs et certains Blancs. Le groupe des ouvriers, par exemple, n'existe pas; un ouvrier noir est d'abord un Noir, et a plus de rapports avec un Noir PDG qu'avec un autre ouvrier blanc.
Distinguer les races humaines, d'un autre côté, la classification linnéenne ne le peut pas. D'abord parce que tous les humains sont interféconds, et qu'il est difficile de maintenir l'idée de transmission héréditaire de l'essence quand deux essences différentes font ensemble un seul rejeton. Ensuite parce que, dans le cas des humains, le racisme est interdit depuis la fin de la Deuxième Guerre mondiale.
La systématique linnéenne bat donc en retraite et se réfugie sur les frontières de l'espèce. Là c'est permis. Nous sommes des humains, eux sont des moutons par exemple, nous avons le droit de les manger. Ils ne sont pas de notre essence. Un fossé infranchissable existe entre eux et nous. Nous, humains, sommes au contraire tous identiques; essentiellement identiques, au moins pour ce qui est des essences que reconnaît la science. Nos différences ne sont pas de l'ordre de la biologie34.
Nous sommes des humains; les chimpanzés sont des chimpanzés. Nous ne sommes pas de la même espèce, nous ne nous reproduisons pas avec eux. Nos parents que nous respectons étaient eux aussi des humains, et les leurs de même. De proche en proche, nos ancêtres sont tous des humains, jusqu'à ce qu'on remonte, par exemple, quelque 300 000 générations en arrière, il y a environ six millions d'années. À ce moment, on trouve quelqu'un qui a la particularité d'être non seulement notre vénérabilissime ancêtre, mais aussi celui des chimpanzés35. Les chimpanzés eux aussi peuvent le réclamer comme leur.
De fait, tous ces gens sont morts. Mais supposons que quelques uns d'entre eux revivent, une poignée répartie au long des deux branches généalogiques qui partent ce vénérable ancêtre commun pour aboutir aux humains d'une part et aux chimpanzés de l'autre. Selon Jared Diamond36, les Indiens d'Amérique se seraient installés sur ce continent il y a environ onze mille ans. Aucun problème d'interfécondité n'a été constaté entre eux et les Européens. Il en est de même sans doute pour d'autres populations37. On peut donc supposer, en extrapolant à partir de cette donnée, qu'il suffirait de faire revivre au maximum six cents ancêtres au total – bien moins suffiraient sans doute38 – pris sur les deux branches généalogiques, pour que nous soyons obligés de classer les humains non dans une espèce, mais à une des deux extrémités d'une espèce en «anneau brisé» comme celle qui regroupe les goélands mentionnés plus haut. À l'autre extrémité il y aurait les chimpanzés.
Les chimpanzés et les humains sont-ils donc de la même espèce ou d'espèces différentes? Ils sont d'espèces différentes, nous dit la systématique; et ceci, non en raison de quoi que ce soit de relatif aux humains, ni de quoi que ce soit de relatif aux chimpanzés, ni même de quoi que ce soit qui se déduise d'une comparaison entre les deux. Ils sont d'espèces différentes parce que les intermédiaires se trouvent être morts.
Mais comment la systématique peut-elle être la systématique, si elle classe en fonction de critères aussi arbitraires, aussi contingents, en fonction de critères qui n'ont rien à voir avec les individus classés? Pire, comment peut-elle prétendre être la systématique scientifique? La seule réponse possible, c'est que l'inexistence de ces intermédiaires est perçue comme non accidentelle. Si c'est un hasard, c'est un «heureux hasard»; c'est la main de la Providence qui nous a séparés des chimpanzés, nous permettant d'être, nous et nous seuls, d'une autre espèce qu'eux, d'être seuls créés à l'image de Dieu, selon son bon Plan39.
D'une certaine façon, pour la systématique linnéenne, ce qui est perçu comme accidentel, c'est l'existence de l'évolution. Le résultat, lui, est essentiel; il correspond au «Plan de la Nature» qu'elle prétend refléter. L'espèce humaine et celle des chimpanzés sont pour elle essentiellement différentes; l'évolution, ce n'est qu'une histoire, l'histoire de comment le Plan a été réalisé.
La race, nous disent les antiracistes, n'est pas un concept scientifique. Et ils ont raison, malgré son apparente «évidence». Mais ils n'ont pas toujours donné les bonnes raisons. Le racisme n'est pas scientifique, disent-ils, puisque les humains se reproduisent tous entre eux40. C'est vrai, dans le cadre de la systématique linnéenne; mais en utilisant pour réfuter le racisme les concepts de la classification linnéenne, ils cautionnent ce même système qui, par sa logique, tend par ailleurs à ressusciter encore et toujours le racisme.
Le racisme n'est pas un concept scientifique d'abord parce qu'il découle de la systématique linnéenne qui, intrinsèquement essentialiste, n'est pas scientifique41. La systématique linnéenne est une menace permanente de retour du racisme42. Il fallait peut-être être antispéciste pour le voir.
La systématique linnéenne est à mettre à la poubelle. C'est là une conclusion un peu forte, diront les biologistes. Commençons donc simplement par cesser de l'appeler la systématique. Qu'est-elle donc alors? Rien; si elle n'est plus la systématique, elle tombe en mille morceaux, et les éléments épars dont elle se prévalait pour justifier ses classifications reprennent leur existence autonome. La morphologie, l'écologie, l'histoire naturelle existent, et peuvent se développer et collaborer, sans l'hégémonie d'une classification. Suivant les besoins, on classera en grands et en petits, en arbres et en herbes, en aquatiques et en terrestres, et, à l'occasion, on classera aussi en descendants ou non du plus proche ancêtre commun des canards et des tortues.
Au niveau de l'espèce, on peut très bien conserver le concept de spéciation, désignant par là le phénomène particulier au monde biologique qui, du fait de la reproduction et de la sexualité, amène à la formation de populations au sein desquelles se pratique l'échange du matériel génétique et qui sont isolées des autres sur ce point. Il ne s'agirait pas d'une classification des individus, mais d'une notion phylogénétique impliquant un grand nombre d'individus. De fait pourtant, la spéciation s'accompagne généralement d'une divergence à un niveau ou un autre entre les groupes; quand un individu appartient à une telle population reproductivement isolée, ce fait est le plus souvent en corrélation avec d'autres de ses caractéristiques. La taille des adultes est généralement moindre chez les chats que chez les lynx. Cette corrélation n'implique nullement cependant de retourner à l'espèce comme critère de classification objective; si c'est à la taille qu'on s'intéresse, c'est la taille qui représente le critère de classification, non l'espèce. De même, dans le domaine éthique, si c'est au fait d'être ou non un individu rationnel qu'on s'intéresse, c'est selon ce critère qu'on classera, et non selon l'espèce, bien que les humains soient généralement, mais pas toujours, plus rationnels que les autres animaux. Et selon le critère de sensibilité, le plus déterminant à mes yeux, on classera sous ce rapport les individus humains embryons avec les plantes.
De fait, d'ailleurs, le degré de corrélation entre espèce et caractère individuel est très variable. Le microbiologiste clinique est tout à fait insatisfait de connaître l'espèce d'un germe, laquelle souvent ne lui apprendra pas si celui-ci est pathogène ou non ou résistant à un antibiotique. À l'inverse, chez les souris par exemple, il existe des «espèces jumelles», qu'on ne sait distinguer que par des analyses biochimiques poussées. C'est que le critère de base de l'espèce – l'interfécondité – correspond, biologiquement, à un pot-pourri de phénomènes hétéroclites. Le fait que deux individus ne puissent se reproduire ensemble peut provenir de leur manque d'attirance sexuelle, d'une incompatibilité morphologique ou comportementale quelconque empêchant l'accouplement, d'une incompatibilité entre la mère et le fœtus, ou d'un problème génétique, métabolique, hormonal ou autre; sans parler des barrières géographiques (présentes et... futures!) ni des changements dans le sens du vent (qui parfois intervient dans la pollinisation). L'interfécondité indirecte dépend aussi, on l'a vu, de l'existence ou non d'individus tiers. L'espèce, qui se veut le résumé de tout, ne représente en réalité rien de déterminé. Elle n'a la valeur cardinale qu'on lui donne que parce qu'elle est la clé du système idéologique qui l'accompagne.
Au niveau de la vie quotidienne, on pourra continuer à appeler un chat un chat, mais aussi une grenouille une grenouille, un ver un ver et un arbre un arbre. Ces trois dernières catégories ne sont pas des espèces reconnues par la classification linnéenne, ni même des regroupements d'espèces. Si on peut appeler un chat un chat, on peut aussi appeler un Noir un Noir, mais Ek par exemple, chat ou pas, je continuerai à l'appeler Ek. Et à souhaiter que vienne le jour où les individus seront simplement ce qu'ils sont, avec leur histoire et leurs histoires, avec leurs désirs et leur vie, sans être «d'abord» quoi que ce soit.
1. Je dirai par la suite simplement «linnéenne», bien qu'elle ait bien sûr évolué depuis Linné.
2. «Taxinomie» veut dire classification. Mais quand en biologie on parle de la taxinomie, c'est à la seule hiérarchie linnéenne qu'on se réfère.
3. En dermatologie c'est même une catégorie tout à fait pertinente. Bien sûr, il s'agit d'une catégorie floue – toutes les gradations existent entre peaux claires et foncées. Mais une roche aussi peut être plus ou moins métamorphique, cela n'invalide pas la classification géologique qu'on peut fonder sur ce caractère.
4. Ce n'est pas le caractère parfois imprécis de ces catégories ou le fait qu'elles dépendent du point de vue adopté qui les rend non scientifiques; en aucune science on n'exige des catégories qu'elles soient parfaitement précises et totalitaires. Ce qu'on ne peut faire, c'est intégrer ces sous-espèces au schéma linnéen.
5. C'est-à-dire, les humains.
6. Hypostasier: considérer comme une substance, attribuer une existence réelle à (dict. Flammarion).
7. Éd. Fayard, 1982, 1989.
8. L'idée de m'intéresser de beaucoup plus près à l'impact que le darwinisme devrait avoir, mais n'a pas eu, sur notre système de pensée m'a été inspirée par le livre de James Rachels, Created from Animals: The Moral Implications of Darwinism, éd. Oxford University Press, Oxford, 1990.
9. Dès le xviiie siècle des géologues avaient commencé à donner à la terre un âge plus avancé; Buffon en 1779 parlait de 168 000 ans (Mayr, p. 306). Le dogme de la création divine subsista cependant presque intact jusqu'à la publication par Darwin en 1858 de L'Origine des espèces, et restait encore vivace parmi les biologistes au début du xxe siècle.
10. Genèse 1:24 par exemple.
11. Individu mâle ou femelle, on ne sait.
12. G.L. Buffon, L'Histoire naturelle, 1749, cité dans Mayr, p. 255.
13. Pages 180 et 181.
14. J. Ray, Hist. Plant., 1686, cité dans Mayr, p. 251.
15. Mayr, p. 257.
16. Buffon, L'Histoire naturelle, cité par Mayr, p. 256.
17. G. Cuvier, lettre à Pfaff, cité dans Mayr, p. 252.
18. En botanique c'est moins le cas qu'en zoologie; si on parle d'«angiospermes» ou de «composés», en fonction d'un caractère «déterminant» du groupe, il y a aussi les «césalpinacées», en souvenir du botaniste Césalpin.
19. La phylogénèse d'un groupe est son histoire évolutive.
20. Mayr, p. 264.
21. Page 265.
22. En donnant une descendance fertile, condition implicite dans l'idée de «perpétuation». Je sous-entends cette condition dans la suite.
23. On pourrait rendre plus précis le concept de «proche parent», ou le laisser flou, cela n'a aucune importance ici. Notons aussi que l'expression «est susceptible de» peut s'interpréter de diverses manières, mais ce n'est pas non plus le sujet.
24. R. Dawkins, «Abîmes dans la pensée», dans The Great Ape Project, ss la dir. de P. Cavalieri et P. Singer, éd. The Fourth Estate, Londres, 1993; trad. G. Ewing et D. Olivier.
25. Puisqu'il y avait des variations accidentelles, on ne pouvait être sûr que celles-ci ne pouvaient jamais elles aussi empêcher la reproduction. Il y avait bien là, mais indépendamment de la question de la fermeture transitive, une difficulté pour l'essentialisme, puisque si deux groupes interféconds devaient bien toujours être de même essence, la réciproque n'était pas nécessairement vraie. On ne pouvait jamais être sûr que deux groupes étaient d'essence différente; peut-être qu'étaient seulement accidentelles les raisons qui empêchent les chats et les chiens de se reproduire entre eux, leur essence étant la même. Et de fait, pour Linné, l'essence se rapportait non à l'espèce mais au genre, c'est-à-dire à un ensemble d'espèces. Cela se retrouve dans le «binôme linnéen», système de dénomination des espèces qu'il inventa et qui forme encore aujourd'hui la base de la nomenclature. Chaque espèce y est désignée par le nom de son genre suivi d'un qualificatif, adjectif par exemple. Le Rat est Rattus norvegicus, c'est-à-dire est une variété de Rattus, un Rattus de variété norvegicus (norvégien). Son essence, ce qu'il est, est Rattus; l'espèce particulière à laquelle il appartient est, elle, vue comme accidentelle. On peut noter d'ailleurs la persistance encore actuelle de tentatives pour établir la notion de genre comme donnée «objective», «naturelle», au contraire des catégories taxinomiques supérieures reconnues comme possédant un certain caractère arbitraire; et que ces tentatives sont, malgré tout, elles aussi fondées sur l'interfécondité (les individus d'espèces différentes mais d'un même genre donneraient une descendance viable, mais non fertile).
Cela ne change rien au fond au lien que j'établis dans cet article entre la systématique linnéenne et l'essentialisme. En ce qui concerne le spécisme et la valeur d'essence attachée au fait d'être humain, notons que l'on dit aussi bien «genre humain» qu' «espèce humaine», et dans la classification linnéenne actuelle le genre Homo n'a qu'une seule espèce, sapiens. On pourrait aussi bien parler de genrisme que de spécisme, il s'agirait en substance de la même chose.
26. On pourrait rendre scientifique la catégorie chat en déterminant l'ensemble des caractéristiques génétiques qu'un individu doit avoir pour pouvoir se reproduire avec au moins un des chats existants. On pourrait alors définir le fait d'être un chat par ces caractéristiques non plus relationnelles mais individuelles. D'une part on notera qu'on n'a jamais tenté une telle réification du concept de chat; c'est que l'entreprise serait loin d'être possible dans l'état actuel de la science, et surtout qu'on considère implicitement le concept de «chat» comme déjà réifié (c'est l'essence «chat»); et d'autre part les espèces ne seraient plus disjointes en théorie: s'il n'existe pas d'animal réel susceptible de se reproduire à la fois avec les chats réels et avec les lynx réels, rien ne dit qu'il ne puisse s'en concevoir. La sauvegarde du principe de séparation des espèces risquerait alors d'impliquer de ne plus classer que dans une seule espèce l'ensemble du monde vivant!
27. Définition citée page 266.
28. Page 275.
29. Par «sexualité» en ce sens on entend non l'existence de deux sexes, mais plus généralement l'échange ou la recombinaison de matériel génétique entre individus différents. Les êtres vivants hermaphrodites, et ceux qui, comme certaines bactéries, échangent des gènes indépendamment de la reproduction, sont sexués en ce sens. Cependant il existe aussi des êtres vivants sans sexualité: certaines plantes par exemple pratiquent l'autofécondation exclusive.
30. Mayr, p. 275.
31. Page 276.
32. Darwin, L'Origine des espèces, cité par Mayr, p. 262.
33. On trouve fréquemment dans la littérature des définitions absurdes du concept de monophylétisme, comme par exemple quand Mayr dit (p. 230) qu'un taxon est monophylétique «quand tous les membres d'un taxon [sont] descendants d'un même plus proche ancêtre commun». Tout groupe d'individus quelconques est alors monophylétique! L'oubli du fait que le groupe doit aussi comprendre tous les descendants de cet ancêtre correspond à mon avis à la préférence de Mayr pour la systématique dite «évolutionniste», dont je parle plus loin.
34. De quel ordre sont-elles donc? La biologie, science de la vie, ne concerne-t-elle pas aussi les humains, êtres vivants? En fait, les différences entre humains ne sont pas vues comme accidentelles non plus, mais comme participant d'un autre ordre encore, le «proprement humain» (ou l'ordre de la liberté).
35. Une des causes de résistance au darwinisme était sans doute qu'on y renversait la séquence hiérarchique traditionnelle qui exige que nous vénérions nos parents, et eux les leurs, et ainsi de suite. Le respect dû aux ancêtres croissait à chaque fois qu'on remonte une génération; alors comment peut-on, à la fin, tomber sur... un singe?
36. J. Diamond, The Rise and Fall of the Third Chimpanzee, éd. Random House, Londres, 1991, pages 100 et 101.
37. Par exemple, les humains qui habitaient la Tasmanie avant la colonisation européenne étaient restés isolés depuis dix mille ans (Diamond, ibid.).
38. On peut suivant le même type d'extrapolation estimer qu'il en faudrait au moins trois au total (un sur chaque branche plus l'ancêtre commun). En effet, deux groupes de chimpanzés existent qui ne se reproduisent pas entre eux, et dont le plus proche ancêtre commun remontrait à il y a environ 2,5 millions d'années (chiffre déduit de Richard Dawkins, article cité). D'un autre côté, les Néanderthaliens, qui se sont séparés de nos ancêtres («hommes de Cro-Magnon») il y a plus de cent trente mille ans, et se sont éteints il y a quarante mille ans, sont classés dans la même espèce que nous. On ne sait cependant pas réellement s'ils étaient interféconds avec les Cro-Magnons (Diamond pense que non, d'autres pensent que si). Prenant cet exemple comme cas limite, l'extrapolation donnerait le chiffre de soixante-dix ancêtres à ressusciter.
39. L'idée de voir potentiellement les humains et les chimpanzés comme une espèce en anneau brisé est tirée de l'article de R. Dawkins cité plus haut. Dawkins, plus que d'autres, semble s'être dégagé de l'emprise de la conception essentialiste de l'espèce. En tant que sociobiologiste, toutefois, il propage à la place une idéologie, qui me semble essentialiste elle aussi, du gène; il n'est pas évident qu'on y gagne toujours.
40. À propos non du racisme mais du sexisme – certains philosophes antiques élevaient aussi les deux sexes au rang d'espèces distinctes – Giulia Sissa nous dit («Philosophies du genre» dans Histoire des femmes en Occident, dir. par G. Duby, t. 1, p. 73): «Dans le langage des sciences contemporaines, il est interdit de dire que le masculin ou le féminin correspondent à une espèce, car l'espèce se définit par la capacité de reproduire des individus qui lui sont conformes (...).» Le verbe «interdire» qu'utilise ici G. Sissa est un peu fort et porte à croire que l'on doit le comprendre au plan non seulement scientifique mais aussi social – justement parce que les espèces, loin d'être de simples objets scientifiques, sont porteuses d'une essence.
41. La systématique linnéenne était une théorie scientifique – bien que fausse – à l'époque où elle explicitait ses fondements théoriques, à savoir la Création divine, les essences et le Plan de la Nature.
42. C'est peut-être en référence inconsciente à cela qu'on appelle parfois le racisme «la bête immonde». Le non-humain, victime de la discrimination spéciste, apparaît comme le support de celle-ci, comme le symbole de la discrimination entre les êtres selon une essence. Mais l'animal ne reste pas en place: la discrimination s'attaque en retour à l'humanité, sous la forme du racisme; la bête est devenue immonde, au lieu de rester tranquillement dans son abattoir.